orange
orange-yellow
bushy
tough
Bahamas
Colombia
Panama
Axinella corrugata







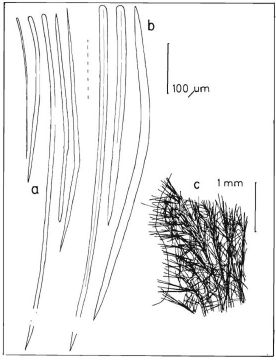
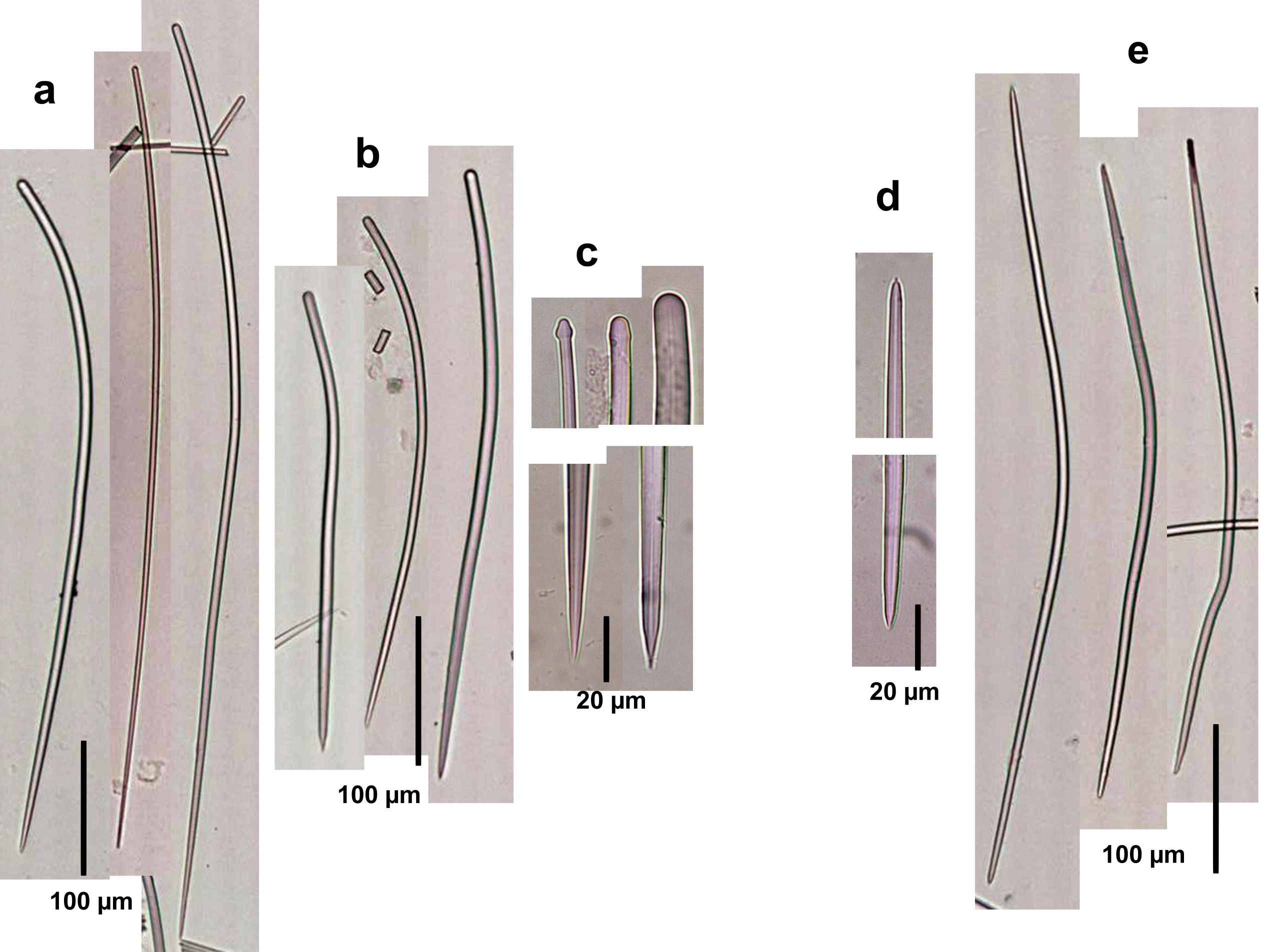
Description: Low bushes, sometimes flattened, up to about 5 m high and wide, with the surface made of flattened or sometimes rounded lamellae or protrusions, often vertically oriented as folds that run vertically for part or the entire sponge. Oscules can be seen in some specimens as small holes (up to about 2 mm) surrounded by a thin membrane. Surface usually velvety, but can be hispid in the ridges of folds. Color orange-yellow. Consistency stiffly compressible. Skeleton as sinuous and confused spicule tracts arising from the center of the bush and ascending and diverging radially towards all surfaces; tracts with 3 to 6 spicules, 40-100 µm thick, embedded in variable amounts of sponging, separated between 100 and 150 µm; scarcely interconnected mostly by single spicules, thus not seen as much as a reticulation. Ascending tracts are more condensed below the surface. The outer surface is hispidated by the terminal spicules of the ascending tracts. Spicules in two types (sizes from measurements from Bahamian and Colombian material, the latter given by Zea, 1987): (1) curved styles, with some heads slightly engrossed terminally or subterminally, and acute points, in two size categories, styles I, 250-442 µm long by 2.3-19µm wide, the most abundant; and styles II, long and slender, but scarcer, 523-974 µm long and 2.4-19 µm wide. (2) Slightly curved oxea with blunt to acute ends, mostly with symmetric ends, 304-807 µm long and 2.4-21.4 µm wide.
Notes: This species lives exposed in the deep reef. It was originally placed under genus Acanthella. It was also known as Oxeostylon (or Teichaxinella) burtoni de Laubenfels, 1934. Initially, we were lumping together all orange to orange-yellow bushy, tubular, subtubular and ramose individuals having two categories of styles and oxea under A. corrugata. Closer inspection led us to separate the material by the overall shape in A. corrugata proper, A. ?digitiformis Lehnert & van Soest, 1996, A. ?pomponiae Alvarez, van Soest & Rützler, 1998, and Axinella sp.-"orange corrugated tubes". (A fifth species with the same spicule complement of the others, Axinella sp.-"orange-yellow velvety tubes", had a shape so different that it was clearly a different species.) In general, A. corrugata has larger styles I and oxea than the othes species, when co-existing specimens are compared. The most similar in shape to A. corrugata is A. ?digitiformis; because of the consistently subtubular habit of the latter we decided to keep it as a separate species until further studies; unfortunately, we did not save material of the latter species for more detailed study of spicules and skeleton.
Molecular studies have found that A. corrugata and some other species of the genus Axinella are not closely related to the type species of the genus Axinella, A. polypoides (Schmidt, 1862), which led to the erection of the phylocode genus Cymbaxinella (family Himerhabdiidae, order Agelasida) to contain them (Gazave et al., 2010; Morrow et al., 2012; Redmond et al., 2013). We prefer to follow the Linnean nomenclatorial system over the phylocode, and thus have decided to maintain the name Axinella for this species while the appropriate genus name (available or new) is found for the clade where this and other species belong.
Teichaxinella morchella Wiedenmayer, 1977, originally described from the Bahamas, was synonymized to A. corrugata by Alvarez et al. (1998). Upon closer and repeated comparisons of material, we believe the former is a different species, also pictured here as Axinella morchella. Axinella morchella has a softer and smoother (lustrous) skin, stretched among ridges. In addition, it has only styles as spicules, in contrast to the additional oxea in A. corrugata. In life, they are unmistakably different in shape, color and texture.
Axinella corrugata has been the subject of numerous studies in chemistry of natural products (e.g., Wilson et al., 1999) and their biotechnological production (e.g., Duckworth et al., 2003). It is interesting to note that the above-mentioned confusion between A. corrugata and A. morchella may have permeated these studies and others in molecular systematics (e.g. Holmes & Blanch, 2007; Gazave et al., 2010; Morrow et al., 2012; Redmond et al., 2013). In several cases the material has come from the Bahamas and most probably it could have been actually A. morchella instead of A. corrugata, the former being much more abundant. Indeed, the most recent 18S Ribosomal DNA tree (Redmond et al., 2013, fig. 12) shows two distinct clades of "Cymbaxinella" corrugata that may correspond to these two species. One clade contains the samples from Bocas del Toro, Panama, and the other contains a sample from the Bahamas and a sample from the Florida Keys (and another of unknown origin; see Redmond et al., 2013, file 019 of the supplemental material). As it appears that A. morchella does not occur in Bocas del Toro and the Southern Caribbean (pers. obs. S.Z.), the samples from this locality are probably A. corrugata; the material from the other clade could thus belong to A. morchella. The vouchers need to be reexamined to determine their real identity. At any rate, the results of Redmond et al. (2013) point that the real A. corrugata does not belong to the genus Axinella proper.
Author Reference: (George & Wilson, 1919)
Link: World Porifera Database
.jpg)